
In questa sezione vorrei spendere alcune parole sulle tecniche di biologia molecolare che vengono utilizzate da qualche anno col fine di chiarire le relazioni di parentela tra le diverse specie. Ovunque nel mondo questo tipo di analisi è diventando rutinario e viene applicato nella sistematica di tutti gli organismi viventi; i funghi non sono un'eccezione. Inoltre alcuni laboratori stanno offrendo un servizio di "sequenziamento" anche a privati cittadini ad un costo divenuto relativamente moderato! E' evidente che maneggiare queso tipo di dati richiede però competenze! Il mio obbiettivo è quello di spiegare, per quanto non sia assolutamente semplice, l'uso di alcuni di questi metodi.
Per quanto riguarda il genere Amanita c'è stato un notevole accumulo di dati (sequenze) negli ultimi anni: le specie studiate non sono più solamente quelle americane o asiatiche.
Mi preme fare una premessa che ritengo fondamentale: le tecniche di biologia molecolare, ad oggi, non sono un'alternativa ai sistemi più "classici" di studio dei funghi come molti immaginano. In realtà sono uno strumento complementare, in alcuni casi estremamente potente e risolutivo. Solo la sinergia tra tutti i metodi disponibili può permettere la comprensione della relazione di parentela tra le popolazioni, le specie e qualsiasi gruppo sovraspecifico. Spero di poter affrontare con una certa chiarezza anche alcune delle problematiche legate all'utilizzo di queste tecniche.

Se una popolazione microbica sta crescendo rapidamente non è sempre facile, al microscopio, individuare cellule sufficientemente simili e, comunque, avremmo spesso la sensazione di trovarci di fronte a qualcosa di molto eterogeneo, almeno per forma e dimensioni. Ma non si tratta di individui appartenenti a specie diverse perché sappiamo con certezza che esse derivano da una unica cellula originaria: in gergo una popolazione di questo tipo è un "clone". Semplicemente, ognuna si divide con dei tempi che via via si sfasano rispetto a quelli delle altre; dato che una cellula cresce, fino a quando non decide di dividersi, si avranno cellule molto più grandi, in procinto di dividersi in due, e cellule piccole appena divise; tra questi estremi vi saranno cellule di dimensioni intermedie.
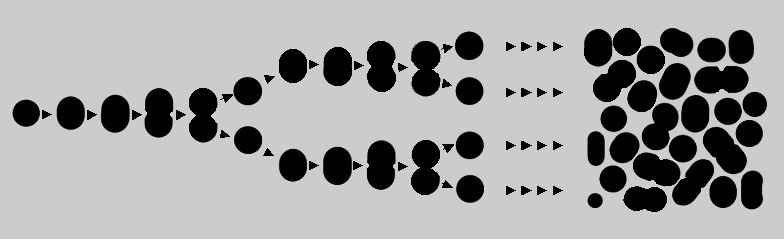
La variabilità può essere causata da diversi fenomeni tra cui una particolare risposta all'ambiente, cioè alle sue diverse componenti come la disponibilità di determinati nutrienti, la presenza o meno di acqua, la composizione chimica del substrato su cui cresce l'organismo, la sua acidità, la temperatura, la luce, ecc., mille variabili difficilmente analizzabili in natura ma più semplicemente controllabili tra le mura di un laboratorio o, meglio, all'interno di una provetta.
I batteri sono organismi molto più semplici dei funghi e, sicuramente, più facilmente studiabili. Eppure anch'essi mostrano un notevole grado di variabilità che, a volte, inganna tanto da rendere difficile la collocazioni tassonomica se non ricorrendo a tecniche molto particolari. I funghi sono organismi più complessi e questo aggiunge un ulteriore livello di variabilità che, però, è più difficile da interpretare, anche in un laboratorio attrezzato. Ciò che mi preme dire è che decidere di proporre una specie nuova basandosi su un certo numero di caratteri dal valore incerto è un operazione che può essere pericolosa. Ho il dubbio che alcuni "studiosi" producano specie nuove più per soddisfare il loro Ego che altro!
Qual'è il criterio con cui si può decidere che un carattere ha un valore tassonomico importante? E quale può essere la sua variabilità per una determinata specie? Problemi, secondo me, difficili da chiarire. L'appassionato tende a dare un grande valore a caratteri piuttosto evidenti ma non necessariamente importanti; per chiarire torniamo all’esempio del colore. E’ evidente per tutti quanto alcune specie, tra cui Amanita muscaria è un esempio noto, siano variabili per la loro colorazione: la variabilità del colore del cappello può essere accompagnata dalla presenza di tonalità gialle più o meno intense in altre parti del carpoforo. Le varianti possono essere presenti nello stesso ambiente, dipendentemente anche dallo stadio di maturazione.

In Amanita muscaria sono presenti alcuni pigmenti che danno colorazioni diverse tra cui giallo, arancio e porpora [H. Musso (1979)]. Basterebbe combinare i pigmenti in rapporti diversi per avere molteplici colorazioni. La quantità di ciascun pigmento potrebbere dipendere da fattori ambientali ma anche dalle caratteristiche peculiari di un singolo sporoforo. Inoltre vi potrebbe essere una causa genetica, una mutazione insorta nello sporoforo, oppure presente già prima, nel micelio. Qual'è il peso del carattere "colore", in una circostanza simile, dal punto di vista tassonomico? Mi sento di rispondere: "irrilevante!"
Le mutazioni sono eventi naturali, sebbene poco frequenti, che possono avere effetti più o meno evidenti; esse dipendono da errori che vengono compiuti durante la replicazione del DNA, anche per opera di agenti mutageni (cioè che causano mutazioni) di varia natura. Tornando al caso del colore immaginiamo che, in una data specie, questo dipenda prevalentemente dalla presenza di un pigmento corrispondente ad una molecola che chiamiamo, per comodità, P. Una molecola può essere introdotta dall'esterno, per esempio con il cibo, oppure viene fabbricata internamente alle cellule attraverso "catene biosintetiche" che possono essere anche molto complesse. Immaginiamo che P venga sintetizzata a partire da un'altra molecola, A, incolore, che viene a sua volta trasformata in altre molecole intermedie, B, C e D, secondo il seguente schema:
Ipotizziamo che D sia una molecola che determina il colore arancio. Ogni passaggio, indicato da una freccia "->", è una reazione chimica che, normalmente, all'interno delle cellule è catalizzata (potremmo dire assistita) da un "enzima" (E nell'esempio sotto riportato) :
A ---> B
Gli enzimi che catalizzano questo tipo di reazioni sono generalmente costituiti da una o più "proteine", molecole estremamente complesse, con la capacità, appunto, di rendere possibili le reazioni chimiche che altrimenti non avrebbero la possibilità di avvenire spontaneamente (non tutte le proteine hanno "attività enzimatica", sia chiaro!). La maggior parte degli enzimi ha una specificità molto alta perché è in grado di catalizzare un solo tipo di reazione, mentre altri producono un certo tipo di modifica su una molecola (il "substrato") ma possono avere substrati relativamente differenti: su ognuna producono quella specifica modifica.
Nel nostro caso lo schema sarà quindi:
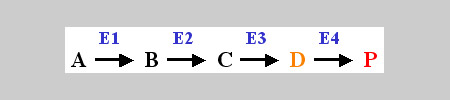

Cosa centrano le mutazioni con gli enzimi? Abbiamo detto che gli enzimi sono generalmente costituiti da proteine; l'informazione per la fabbricazione di una proteina risiede all'interno di un gene, ossia una porzione specifica della lunga catena di DNA (contenuta in ciascuna cellula); le mutazioni sono variazioni della struttura chimica del DNA stesso: se un gene viene modificato l'informazione contenuta in esso può cambiare; a questo cambiamento consegue anche quello della proteina che potrebbe funzionare oppure no dipendentemente dall'entità del cambiamento: alcune mutazioni non hanno alcun effetto mentre altre hanno un effetto deleterio. Altre mutazioni possono avere un effetto intermedio tra i due estremi. L'effetto deleterio di una mutazione può determinare nei casi estremi l'incompatibilità con la vita. Le mutazioni sono però la base di cambiamenti che, se ereditati, permettono di accumulare caratteri nuovi che, in tempi lunghi, causano incompatibilità tra popolazioni di individui causando la nascità di nuove entità (per esempio nuove specie).


In realtà una parte o tutto il pigmento D potrebbe essere convertito in P a meno che anche E4 non sia cambiato contemporaneamente.
E se fossero E1 o E2 ad avere dei problemi? Se la funzione fosse solamente ridotta probabilmente si verificherebbe anche in questo caso un accumulo più o meno limitato di D e P dando un colore più o meno chiaro . E' ovvio che se E1 (da A a B) oppure E2 (da B a C) o E3 (da C a D) non funzionassero per niente si otterrebbero probabilmente degli sporofori albini, completamente depigmentati.
Inoltre se la distribuzione della pigmentazione non fosse omogenea e se i problemi di sintesi fossero localizzati solo in certe porzioni, indipendenti dal resto, si avrebbe un'ulteriore complessità. Si potrebbe giocare con i colori all'infinito.
E quanto possono influire le caratteristiche ambientali?
Un'unica cosa mi sentirei di dire a questo punto: è assolutamente inutile e fuorviante continuare a creare forme e varietà che altro non sono che un'espressione della variabilità biologica o, a volte, della psicologia del micologo! Chi frequenta ambienti diversi sa quanto spesso le specie cambino il proprio aspetto. Si potrebbe obiettare: e se fossero effettivamente specie diverse?
Torniamo ad una delle domande precedenti: qual'è il criterio con cui si può decidere che un carattere ha un valore tassonomico importante? E quale può essere la sua variabilità per una determinata specie prima di poter affermare che ciò che abbiamo davanti agli occhi sia qualcosa di realmente diverso dalla specie nota che avevamo immaginato in prima istanza?
Un carattere con valore tassonomico "importante" non dovrebbe essere soggetto a variazioni causate da motivi banali, come potrebbe essere il colore. La struttura della volva, ad esempio, dipende sicuramente da un certo numero di fattori che interagiscono in modo complesso tra di loro sin dal livello molecolare: tutto ciò si riflette nella struttura cellulare, visibile al microscopio, da cui dipende, poi, l'aspetto macroscopico. Differenze di una certa consistenza in questo caso probabilmente riflettono una reale distanza evolutiva proprio perché se molti fattori devono cambiare per avere modifiche di certi caratteri sono necessarie molte mutazioni e, di conseguenza, tempi lunghi (e, probabilmente, mutazioni in parecchi geni da cui potrebbe derivare un'incompatibilità tra i diversi genomi (mi fermo!).